Биологични молекулярни машини
Една интересна тема в контекста на съвременните постижения в молекулярната биология. Тази научна дисциплина се фокусира върху разбирането на живите организми на молекуларно ниво и разкриването на основните механизми, които управляват жизнените процеси. Откритията в молекулярната клетъчна биология през последните десетиления промениха начина, по който разбираме живота и функциите на живите организми. Революцията в молекулярната биология е безпрецедентна по своя мащаб и се отразяват в живота на всеки от нас.
Въпросът за биологичните машини може да бъде разглеждан от различни гледни точки, което дава възможност за интересни дебати в областта на науката и философията. Някои учени използват термина "молекулярни машини" в своите научноизследователски документи за да опишат сложните биологични процеси и функции в клетките. Този термин може да се разглежда като удобна аналогия, която ни помага да си представим сложността на биологичните системи и тяхната способност да изпълняват специфични задачи, подобно на създадените от човека машини. От друга страна, има учени и научни философи, които подчертават уникалността на биологичните системи и не смятат, че аналогията с машини е напълно подходяща. Те смятат, че организмите в хода на еволюцията са развили множество сложни комплекси, които се различават от машините, създадени от човека.
Традиционно терминът "машина" се свързва с устройство, което извършва механична работа за изпълнение на специфични задачи. Машина (на старогръцки: Μηχανή; на латински: machina (-ae, f.)) е сложен механизъм или съчетание от механизми, който изпълнява механични движения за преобразуване на енергия и материали, за извършване на работа, за събиране, предаване, съхранение, обработка и използване на информация.
Въпреки това, съвременното развитие в технологиите и науката предизвика разширяване на концепцията за машини. В днешно време, машините са още по-разнообразни и може да се отнасят и до устройства, които не извършват само механична работа. Така че, в съвременния свят, терминът "машина" може да обхваща различни устройства и системи, които изпълняват специфични задачи, независимо дали са механични, електронни, биологични или други. Това е следствие на продължаващият технологичен напредък и научни открития.
Според определението за машина, клетъчните структури, които можем да определим като "машини", са тези, които изпълняват специфични функции чрез механични движения, подобно на това как работят изкуствените машини, които са създадени от човека. Всъщност, списъкът с биологични машини е много дълъг и разнообразен, ето няколко примера:
- Най-атрактивни са биологичните ротационни машини архелум (Archaellum), флагелум (Flagellum) и АТФ-синтаза (ATP synthase). Има още една ротационна машина - T9SS, която на този етап е слабо проучена.
- След тях се нареждат моторните протеини кинезин, динеин и миозин, отговорни за механични движения и транспорт на структури и органели в клетката.
- Следващите машини влизат в състава на генетичният апарат, това са рибозоми, сплайсозоми, хеликази, ДНК-полимерази, РНК-полимерази, аминоацил-тРНК синтетази (aaRSs) и др.
- Наномашините за секреция тип T1SS до T9SS. Някои от тях (T3SS, T4SS и T6SS) имат поразителна прилика с медицинска спринцовка с игла и цилиндър. Секреционната система тип III (T3SS) наричана инжектизома служи за инжектиране на токсини и ефекторни протеини в целевите гостоприемни клетки. Тя е елемент от факторите на вирулентност в много патогенни бактерии, включително причинителите на чума, холера, салмонела, легионерска болест, коремен тиф, магарешка кашлица, инфекции, предавани по полов път и др. Наноспринцовката (T3SS) на чумния патоген Yersinia pestis е отговорна за големите чумни епидемии в човешката история.
- Всички видове йонни помпи на K+, Na+, Mg2+, Zn2+, Cu2+, Ca2+, Mn2+, Fe2+ и мн. др. йони, задвижвани с протони, натриеви йони или АТФ.
- Помпи задвижвани от светлина - хлоридната помпа халородопсин, протонните помпи бактериородопсин и археродопсин.
- Всички селективни и комбинирани йонни канали, механочувствителни канали, те също извършват механични движения.
- Всички мембранни синпортери и антипортери, транспортери на молекули и субстрати, суперсемейството на касетните АВС-транспортери, семейство SLC25 митохондриални трансмембранни преносители, преносители като TAPL, EmrE, TRAP, MATs, SecY, YidC и мн. др.
- Всички ензими също са машини.
Това е част от разнообразието от молекулярни машини, които работят в клетките, за да осигурят нормалното функциониране на клетъчния живот.
Tерминът "молекулярни машини" се използва от изследователите, когато описват и анализират сложни и прецизни структури и процеси в живите клетки, които изпълняват специфични функции, подобни на машини, но работещи на молекулярно ниво.
Ето само няколко научноизследователски статии в които терминът „молекулярнa машина“ е застъпен в самото заглавие. Съществуват стотици подобни статии, а също така и хиляди други в които терминът машина не пресъства в заглавието, но присъства в статията.
Научноизследователските статиите са публикувани в nature, cell, annualreviews, sciencedirect, frontier, phys.org, mdpi, ncbi, eLife, Elsevier, Wiley и др.
Molecular machines stimulate intercellular calcium waves and cause muscle contraction
(Молекулярните машини стимулират междуклетъчните калциеви вълни и предизвикват мускулна контракция)
https://www.nature.com/articles/s41565-023-01436-w
An early origin of iron–sulfur cluster biosynthesis machineries before Earth oxygenation
(Ранен произход на машините за биосинтеза на клъстери желязо-сяра преди оксигенацията на Земята)
https://www.nature.com/articles/s41559-022-01857-1
Structural variability, coordination and adaptation of a native photosynthetic machinery
(Структурна променливост, координация и адаптация на нативна фотосинтетична машина)
https://www.nature.com/articles/s41477-020-0694-3
A look under the hood of the machine that makes cilia beat
(Поглед под капака на машината, която кара ресничките да бият)
https://www.nature.com/articles/s41594-022-00778-8
Bacterial motility: machinery and mechanisms
(Бактериална подвижност: машини и механизми)
https://www.nature.com/articles/s41579-021-00626-4
Structural properties of [2Fe-2S] ISCA2-IBA57: a complex of the mitochondrial iron-sulfur cluster assembly machinery
(Структурни свойства на [2Fe-2S] ISCA2-IBA57: комплекс от митохондриална машина за сглобяване на клъстери желязо-сяра)
https://www.nature.com/articles/s41598-019-55313-5
Molecular interplay of an assembly machinery for nitrous oxide reductase
(Молекулно взаимодействие на машина за сглобяване на азотен оксид редуктаза)
https://www.nature.com/articles/s41586-022-05015-2
The Hsc70 disaggregation machinery removes monomer units directly from α-synuclein fibril ends
(Машината за дезагрегиране на Hsc70 премахва мономерните единици директно от краищата на α-синуклеиновите фибрили)
https://www.nature.com/articles/s41467-021-25966-w
The Viral Class II Membrane Fusion Machinery: Divergent Evolution from an Ancestral Heterodimer
(Машината за сливане на мембрана от вирусен клас II: Дивергентна еволюция от предшественик хетеродимер)
https://www.mdpi.com/1999-4915/13/12/2368
Lateral opening in the intact β-barrel assembly machinery captured by cryo-EM
(Страничен отвор в непокътнатата машина за сглобяване на β-цевта, уловен от крио-ЕМ)
https://www.nature.com/articles/ncomms12865
Dynamic action of the Sec machinery during initiation, protein translocation and termination
(Динамично действие на машината Sec по време на иницииране, транслокация на протеини и прекратяване)
https://elifesciences.org/articles/35112
Lactose permease: A beautiful chemiosmotic machine
(Лактозна пермеаза: Красива хемиосмотична машина)
https://www.sciencedirect.com/science/article/pii/S0005272812001971?via%3Dihub
The mercury resistance operon: from an origin in a geothermal environment to an efficient detoxification machine
(Оперонът за устойчивост на живак: от произход в геотермална среда до ефективна машина за детоксикация)
https://www.frontiersin.org/articles/10.3389/fmicb.2012.00349/full
AAA+ Proteases: ATP-Fueled Machines of Protein Destruction
(AAA+ протеази: ATP-захранвани машини за разрушаване на протеини)
https://www.annualreviews.org/doi/10.1146/annurev-biochem-060408-172623
The RNA Degradosome of Escherichia coli: An mRNA-Degrading Machine Assembled on RNase E
(РНК деградозома на Escherichia coli : машина за разграждане на иРНК, сглобена на РНаза Е)
https://www.annualreviews.org/doi/10.1146/annurev.micro.61.080706.093440
Apoptosome: The Seven-Spoked Death Machine
(Апоптозома : смъртоносна машината със седем спици)
https://www.sciencedirect.com/science/article/pii/S1534580702001375
The Centromere: Chromatin Foundation for the Kinetochore Machinery
(Центромер: Хроматинова основа за кинетохорната машина)
https://www.ncbi.nlm.nih.gov/pmc/articles/PMC4160344/
Protein-Injection Machines in Bacteria
(Машини за инжектиране на протеини в бактерии)
https://www.cell.com/cell/fulltext/S0092-8674(18)30111-9?_returnURL=https%3A%2F%2Flinkinghub.elsevier.com%2Fretrieve%2Fpii%2FS0092867418301119%3Fshowall%3Dtrue
Inventing the dynamo machine: the evolution of the F-type and V-type ATPases
(Изобретяване на динамо машината: еволюцията на АТФазите от F-тип и V-тип)
https://www.nature.com/articles/nrmicro1767
THE ATP SYNTHASE—A SPLENDID MOLECULAR MACHINE
(АТФ СИНТАЗАТА — ПРЕКРАСНА МОЛЕКУЛНА МАШИНА)
https://www.annualreviews.org/doi/10.1146/annurev.biochem.66.1.717
Structures of LRP2 reveal a molecular machine for endocytosis
(Структурите на LRP2 разкриват молекулярна машина за ендоцитоза)
https://www.sciencedirect.com/science/article/abs/pii/S0092867423000466
The machinery of mitosis: Kinetechores, centrioles and chromosome pumps
(Машината на митозата: Кинетехори, центриоли и хромозомни помпи)
https://phys.org/news/2013-10-machinery-mitosis-kinetechores-centrioles-chromosome.html
Photomechanical molecular machines enable control of cell signalling
(Фотомеханичните молекулярни машини позволяват контрол на клетъчното сигнализиране)
https://www.nature.com/articles/s41565-023-01437-9
Propulsive nanomachines: the convergent evolution of archaella, flagella and cilia
(Пропулсивни наномашини: конвергентна еволюция на архела, флагела и реснички)
https://academic.oup.com/femsre/article/44/3/253/5800988
Hybrid Quantum Mechanical/Molecular Mechanical Methods For Studying Energy Transduction in Biomolecular Machines
(Хибридни квантово-механични/молекулярно-механични методи за изследване на енергийната трансдукция в биомолекулярни машини)
https://www.annualreviews.org/doi/full/10.1146/annurev-biophys-111622-091140
Structure and Mechanism of the Hsp90 Molecular Chaperone Machinery
(Структура и механизъм на машината за молекулярни шаперони на Hsp90)
https://www.annualreviews.org/doi/10.1146/annurev.biochem.75.103004.142738
Photomechanical molecular machines enable control of cell signalling
(Фотомеханичните молекулярни машини позволяват контрол на клетъчното сигнализиране)
https://www.nature.com/articles/s41565-023-01437-9
Bringing Dynamic Molecular Machines into Focus by Methyl-TROSY NMR
(Поставяне на фокус върху динамичните молекулярни машини чрез Methyl-TROSY NMR)
https://www.annualreviews.org/doi/full/10.1146/annurev-biochem-060713-035829
Structure of the Hsp110:Hsc70 Nucleotide Exchange Machine
(Структура на машината за обмен на нуклеотиди Hsp110:Hsc70)
https://www.cell.com/trends/neurosciences/fulltext/S1097-2765(08)00339-0
The Hsp70 and Hsp60 Chaperone Machines
(Машините Hsp70 и Hsp60 шаперони)
https://www.cell.com/fulltext/S0092-8674(00)80928-9?large_figure=true
Molecular machines governing exocytosis of synaptic vesicles
(Молекулярни машини, управляващи екзоцитозата на синаптичните везикули)
https://www.nature.com/articles/nature11320
In Situ Molecular Architecture of the Salmonella Type III Secretion Machine
(In situ молекулярна архитектура на машината за секреция тип III на Salmonella)
https://www.sciencedirect.com/science/article/pii/S0092867417302003?via%3Dihub
Coordinating assembly of a bacterial macromolecular machine
(Координирано сглобяване на бактериална макромолекулна машина)
https://www.ncbi.nlm.nih.gov/pmc/articles/PMC5963726/
The 26S Proteasome: A Molecular Machine Designed for Controlled Proteolysis
(26S протеазома: молекулярна машина, проектирана за контролирана протеолиза)
https://www.annualreviews.org/doi/full/10.1146/annurev.biochem.68.1.1015
Centromeric Heterochromatin: The Primordial Segregation Machine
(Центромерен хетерохроматин: Първичната машина за сегрегация)
https://www.annualreviews.org/doi/full/10.1146/annurev-genet-120213-092033
Single-Molecule Studies of the Neuronal SNARE Fusion Machinery
(Изследвания на една молекула на машината за синтез на невронни SNARE)
https://www.annualreviews.org/doi/full/10.1146/annurev.biochem.77.070306.103621
The molecular machinery of regulated cell death
(Молекулярната машина за регулирана клетъчна смърт)
https://www.nature.com/articles/s41422-019-0164-5
Autophagy as a modulator of cell death machinery
(Автофагията като модулатор на машината за клетъчна смърт)
https://www.nature.com/articles/s41419-020-2724-5
Computational Approaches to Detect Allosteric Pathways in Transmembrane Molecular Machines
(Изчислителни подходи за откриване на алостерични пътища в трансмембранни молекулярни машини)
https://www.ncbi.nlm.nih.gov/pmc/articles/PMC4877268/
Large T Antigens of Polyomaviruses: Amazing Molecular Machines
(Големи Т антигени на полиомавируси: невероятни молекулярни машини)
https://www.annualreviews.org/doi/10.1146/annurev-micro-092611-150154
Кратък преглед на реверсивните ротационни машини Archaellum (в археите) и Flagellum (в бактериите)
Плувната подвижност е полезна за оцеляването на клетките, тъй като позволява промяната от неблагоприятна среда към по-благоприятна в търсене на храна. Архелум и флагелум са два различни типа молекулярни ротационни двигатели, които имат функционално сходство в тяхната основна роля за създаване на движение на клетката чрез въртене на спирално витло (камшиче). Въпреки че и двете структури на подвижност споделят една и съща функция, те са структурно несвързани. Архелумите са съставени от около 10-15 различни протеина, докато флагелумите от около 50. И двата двигателя се въртят както по посока на часовниковата стрелка, така и обратно на часовниковата стрелка (реверсивна функция). При Archaea и Bacteria отделните субединици на въртящия филамент са посттранслационно модифицирани чрез гликозилиране. Гликаните се добавят последователно към хидроксилната кислородна група на серин и треонин.
Археларен двигател
Архелума е изграден от твърда нишка (камшиче) наричана архела с диаметър ∼10 nm и дължина до ~10 μm. Архелата съдържа ~61 500 архелинови субединици. Архелин мономерът с форма на попова лъжица има два домена, β-цевичен домен и дълга, леко извита α-спирална опашка. Всяки един мономер е изграден от 163 аминокиселини. Камшичето се върти от молекулярен реверсивен двигател с диаметър 30 μm вграден в мембраната на клетката. Той се задвижва от хидролиза на АТФ, наблюденията потвърждават необходимост от 12 молекули АТФ за един оборот. Въртящият момент на двигателя е около 160 pN nm при честота 25 Hz, която съответсва на 1500 об/мин. Интересно е да се отбели, че археларният въртящ момент остава постоянен 160 pN·nm независимо от скоростите на въртене между 0,5 и 30 Hz. Промяна на посоката на въртене е сложен и не добре изучен процес. Извстно е участието на пртеин CheY, който предизвиква конформационна промяна в ротора на двигетеля. Превключването на посоката на въртене е по-добре проучено при флагелум.
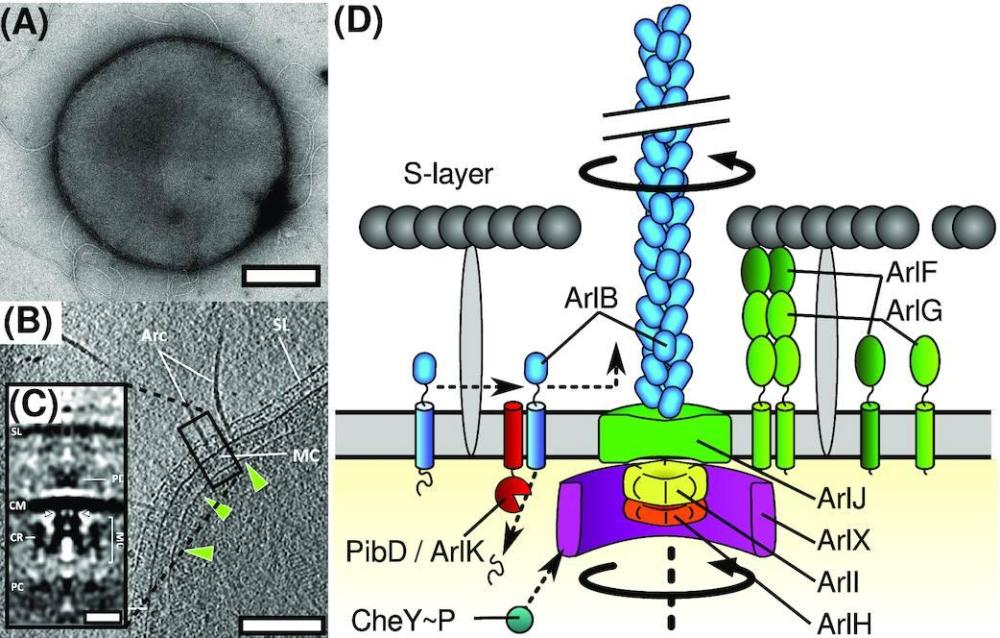
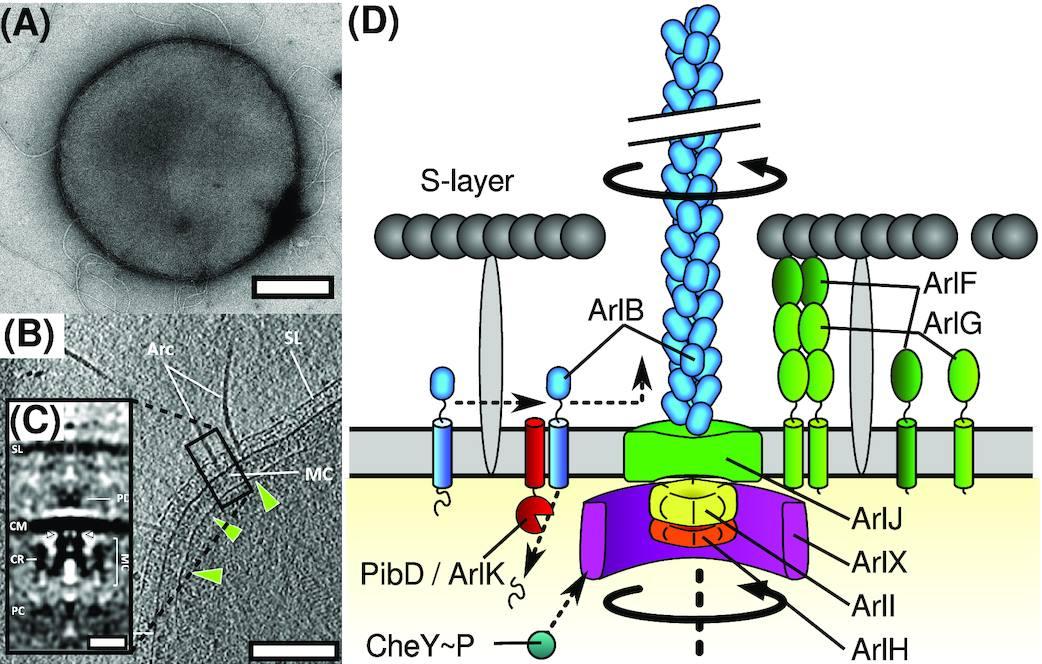
Структура на Archaellum. (A) , Отрицателно оцветена трансмисионна електронна микрофотография на архелирания кренархеон Sulfolobus acidocaldarius , моделен организъм за изследване на архела. Бар, 500 nm. (B) , Томографски срез на клетка Pyrococcus furiosus от Daum et al . 2017 г. , показваща полярна шапка (зелени върхове на стрели), археларни двигатели (или моторен комплекс, MC), S-слой (SL), цитоплазмена мембрана (CM) и архаеларни нишки (Arc). Мащабната лента е 200 nm. (C) , Средна субтомограма на Pyrococcus furiosus archaellar мотор от Daum et al . 2017 гпоказващи периплазмени плътности (PD) и цитоплазмен пръстен (CR). Мащабната лента е 20 nm. (D) , Карикатурна схема на това, което е известно за архитектурата на archaellum.
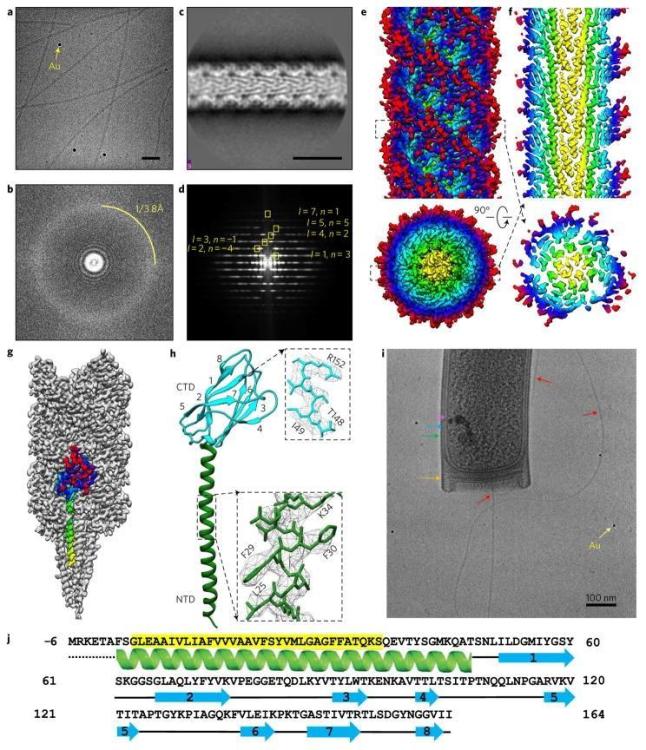
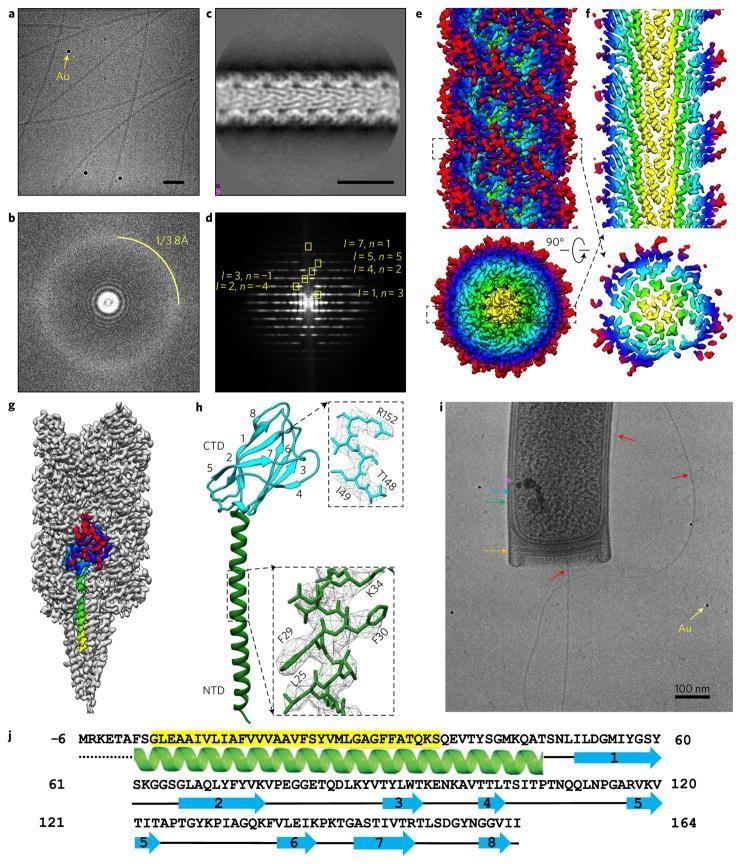
a , Представителна cryoEM микроснимка на нишки от Methanospirillum hungatei JF1 с добавени 10 nm златни флудициали (жълта стрелка). Скала, 50 nm. Около 3300 видеоклипа бяха събрани в набора от данни, от които ~75% имаха влакна, използвани в двуизмерна класификация. b , трансформация на Фурие на микроснимката в a, с указан воден пръстен 3,8 Å. c , Резултат от двуизмерна класификация на CTF коригирани изображения за нишките M. hungatei archaella. Единадесет двуизмерни класа (от които класът на снимката е един) бяха използвани в крайната реконструкция. Скала, 10 nm. d , Преобразуване на Фурие на двумерната средна класа със спирално индексиране. е, Реконструкция на M. hungatei archaella при разделителна способност 3,4 Å. f , реконструкцията, показваща централни α-спирали. g , реконструкция на Archaella с подчертана единична субединица (дъга). h , Реконструкция на архелин с два домена, хидрофобна N-терминална α-спирала (зелена) и C-терминална осемверижна β-барел (циан) с номерирани нишки. Вмъквания: близки планове на α-спиралата и на нишка 7 на β-цевта. i , CryoEM изображение на клетъчен край на M. hungatei с цитоплазмена мембрана (розово), S-слой (син), обвивка (зелено), запушалка (оранжево), 10 nm златен фидуциал (жълт) и изпъкнал архелум (червен), обозначен с стрелки. Скала, 100 nm. й, Схема на първичните и вторичните последователности на протеина M. hungatei FlaB3 (Mhun_3140). Маркираната в жълто част от последователността е запазена при архелините.
Използвани документи :
Нишка от архелум, съставена от две редуващи се субединици
https://www.nature.com/articles/s41467-022-28337-1
CryoEM структурата на Methanospirillum hungatei archaellum разкрива структурни характеристики, различни от бактериалния флагел и тип IV пили
https://www.ncbi.nlm.nih.gov/pmc/articles/PMC5695567/
Измерването на въртящия момент на двигателя на Halobacterium salinarum archaellar предлага общ модел за ATP-задвижвани ротационни двигатели
https://www.nature.com/articles/s42003-019-0422-6
Структурни прозрения в механизма на археларното ротационно превключване
https://www.nature.com/articles/s41467-022-30358-9
Молекулярен анализ на кренархеалния флагелум
https://onlinelibrary.wiley.com/doi/10.1111/j.1365-2958.2011.07916.x
Флагеларен двигател
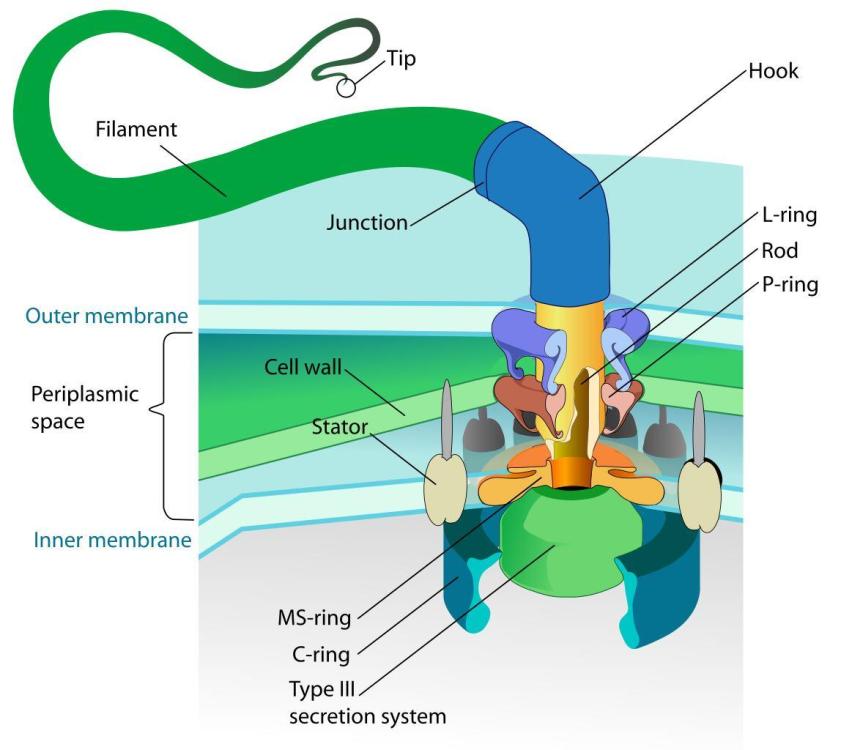
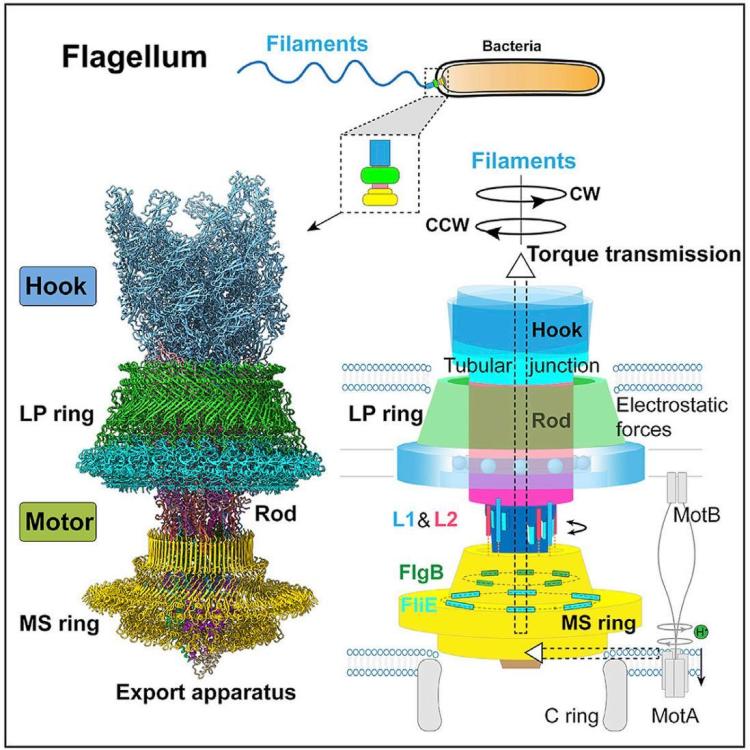
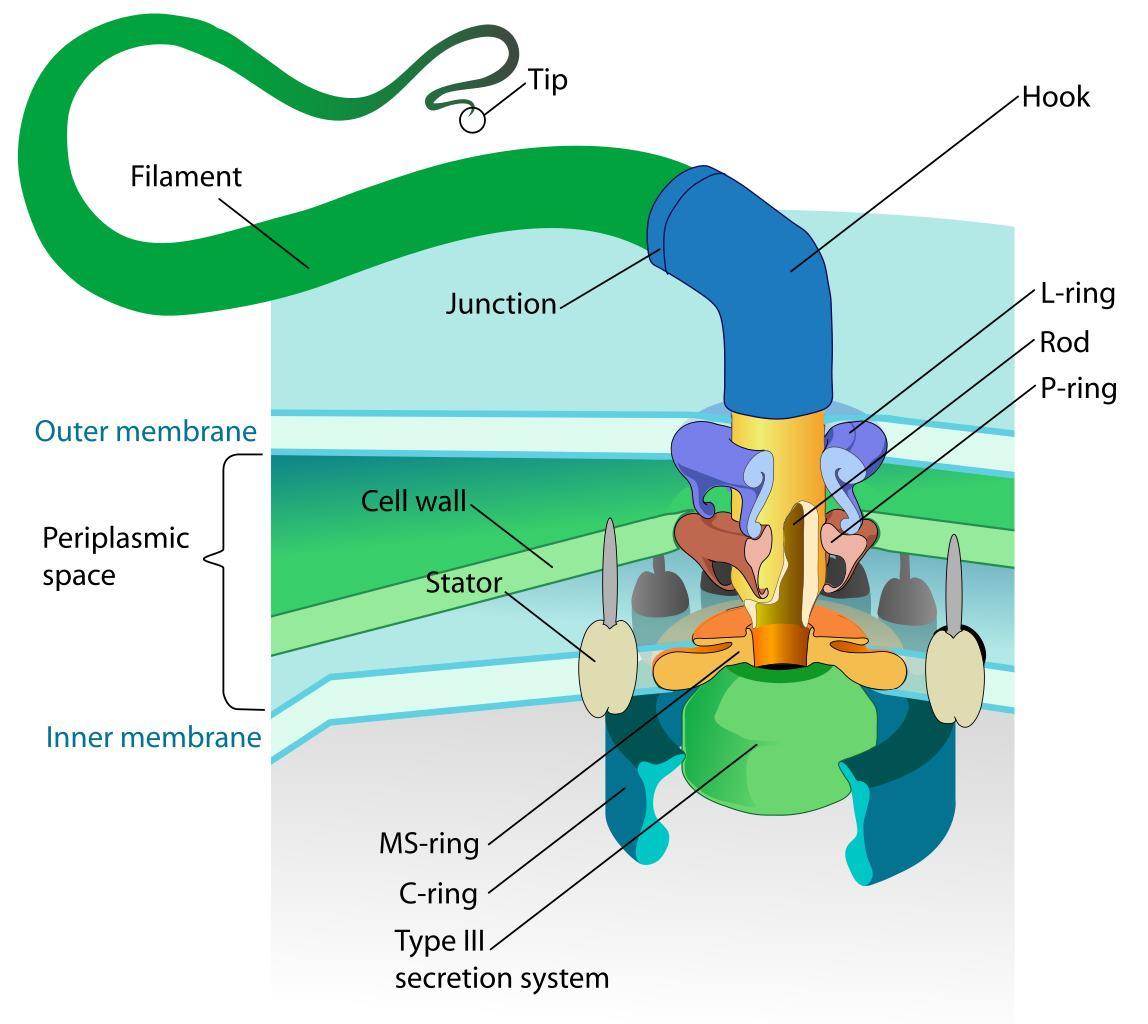
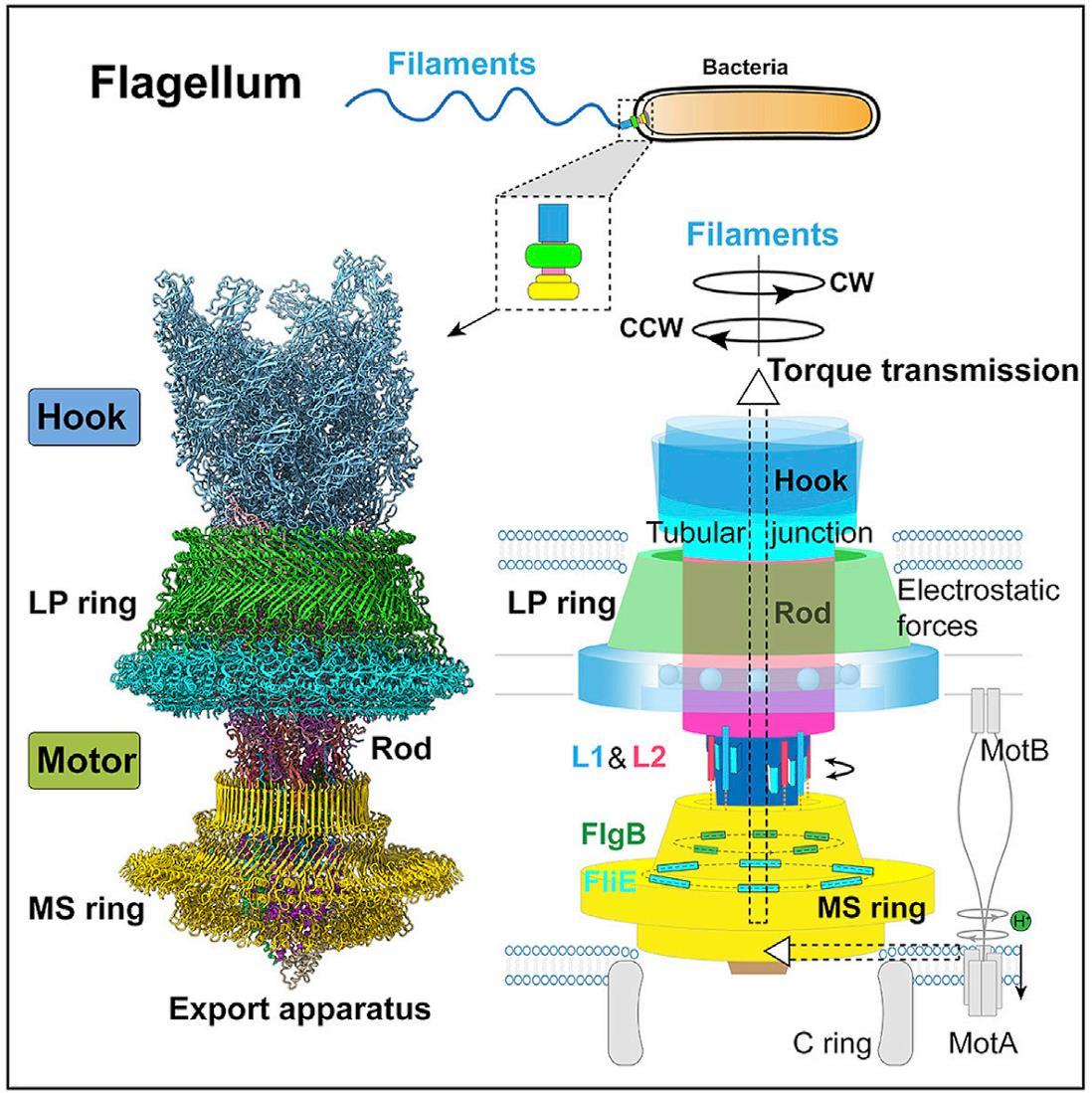
Един от най-очарователните и сложни биологични ротационни двигатели е флагеларният апарат, използван от бактериите, за да се задвижват през течни среди.
Бактериалния флагелум представлява многофункционална и сложна структура, състояща се от няколко ключови компоненти: Флагеларна нишка наподобяваща камшик, която е извън бактерията и изпълнява ролята на винт; ротационен реверсивен двигател състоящ се от статор, ротор и лагери. Двигателят се задвижва от протонна движеща сила (pmf) или натриеви йони (Na+) през мембраната на клетката. Флагелумът е изключително ефективен молекулярен двигател, който има впечатляващи характеристики. Ротора се върти се с честота f =300–1200 Hz, което отговаря на 18 000 до 72 000 об/мин и въртящ момент 1200 pN nm. Максималният въртящ момент на BFM в E. coli е ∼2000 pN nm. Моторът на C. jejuni има три допълнителни диска, отчитайки въртящия момент от ∼3600 pN nm. При тези обороти бактериите могат да се придвижват със скорост от 30 μm/s.
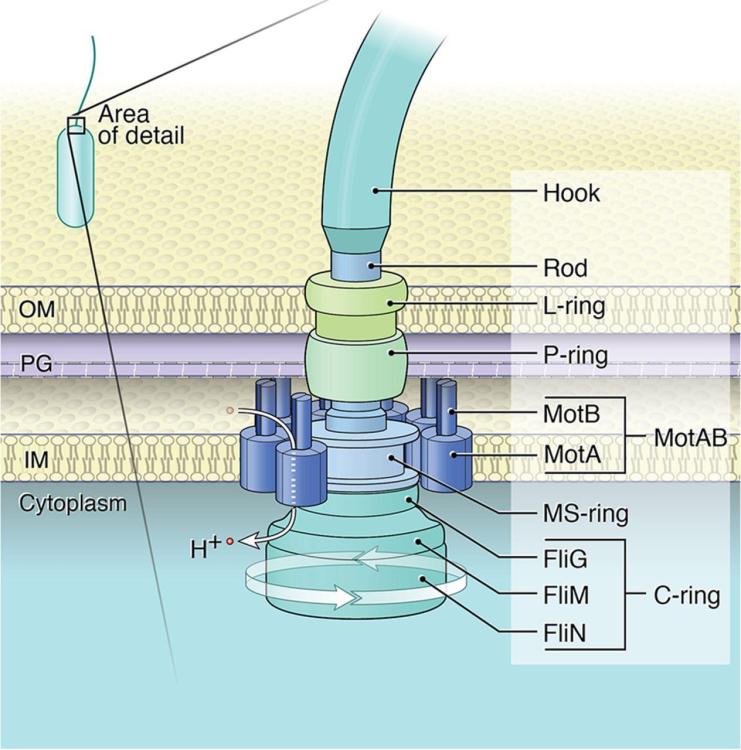
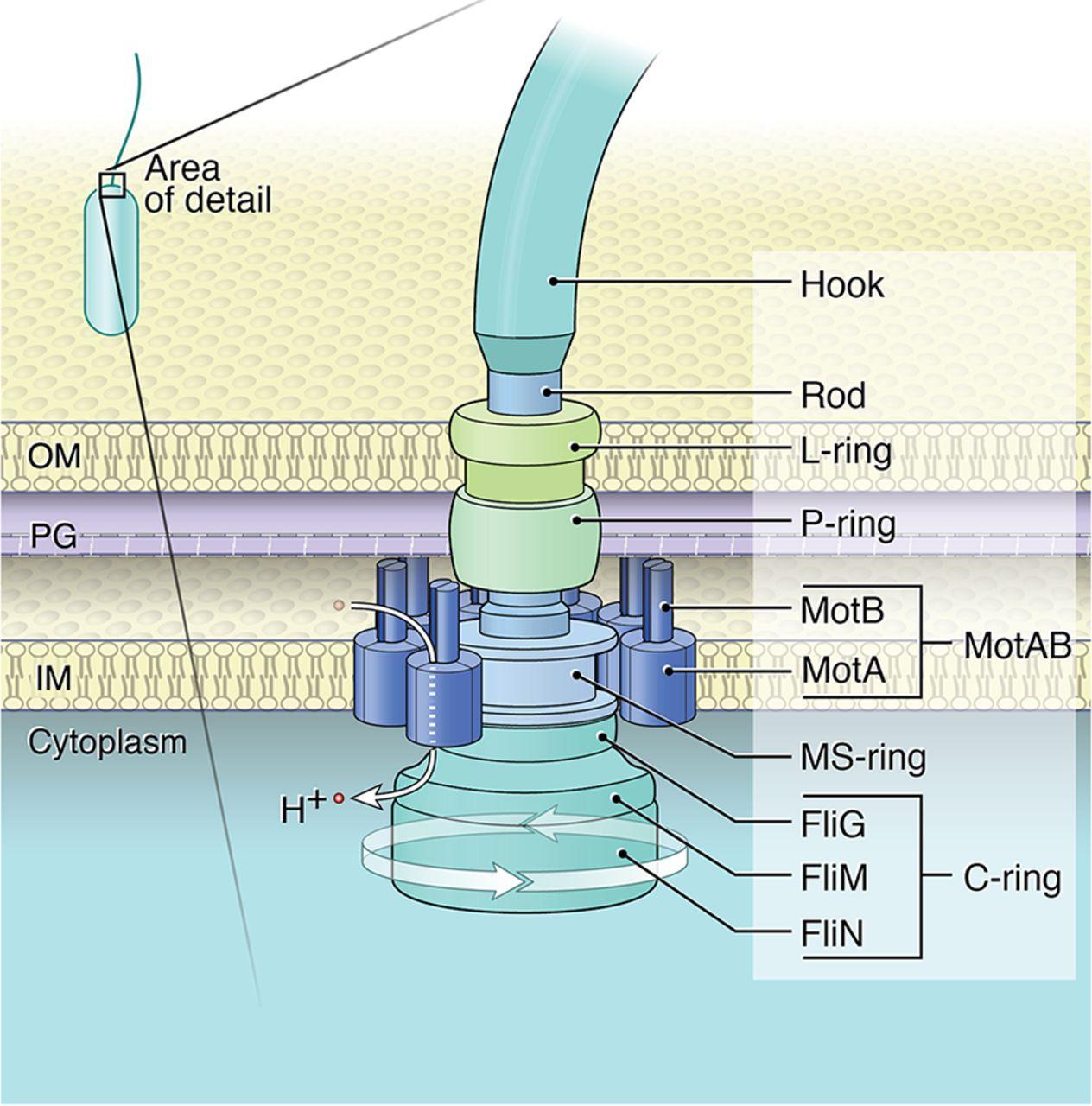
Свързаният с мембрана статор и превключващият комплекс (наричан още С-пръстен) са пряко отговорни за въртенето и превключването на флагеларите. Статорният комплекс е генератор на въртящ момент, захранван от йонен поток през мембраната и се състои от няколко статорни единици, фиксирани в мембраната
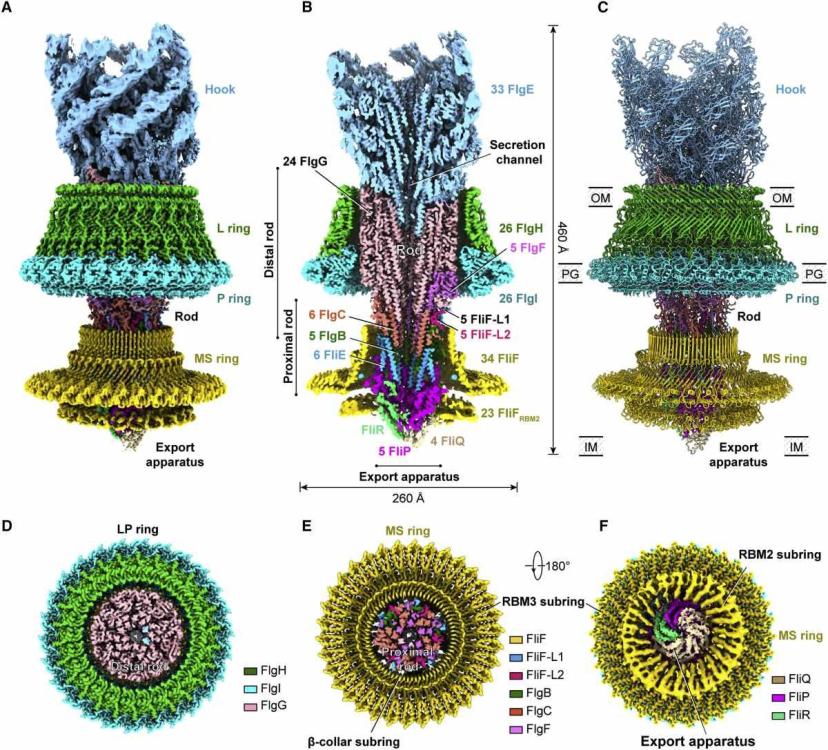
Cryo-EM структура на бактериалния флагеларен двигател в комплекс куката
Обща структура на флагеларния комплекс
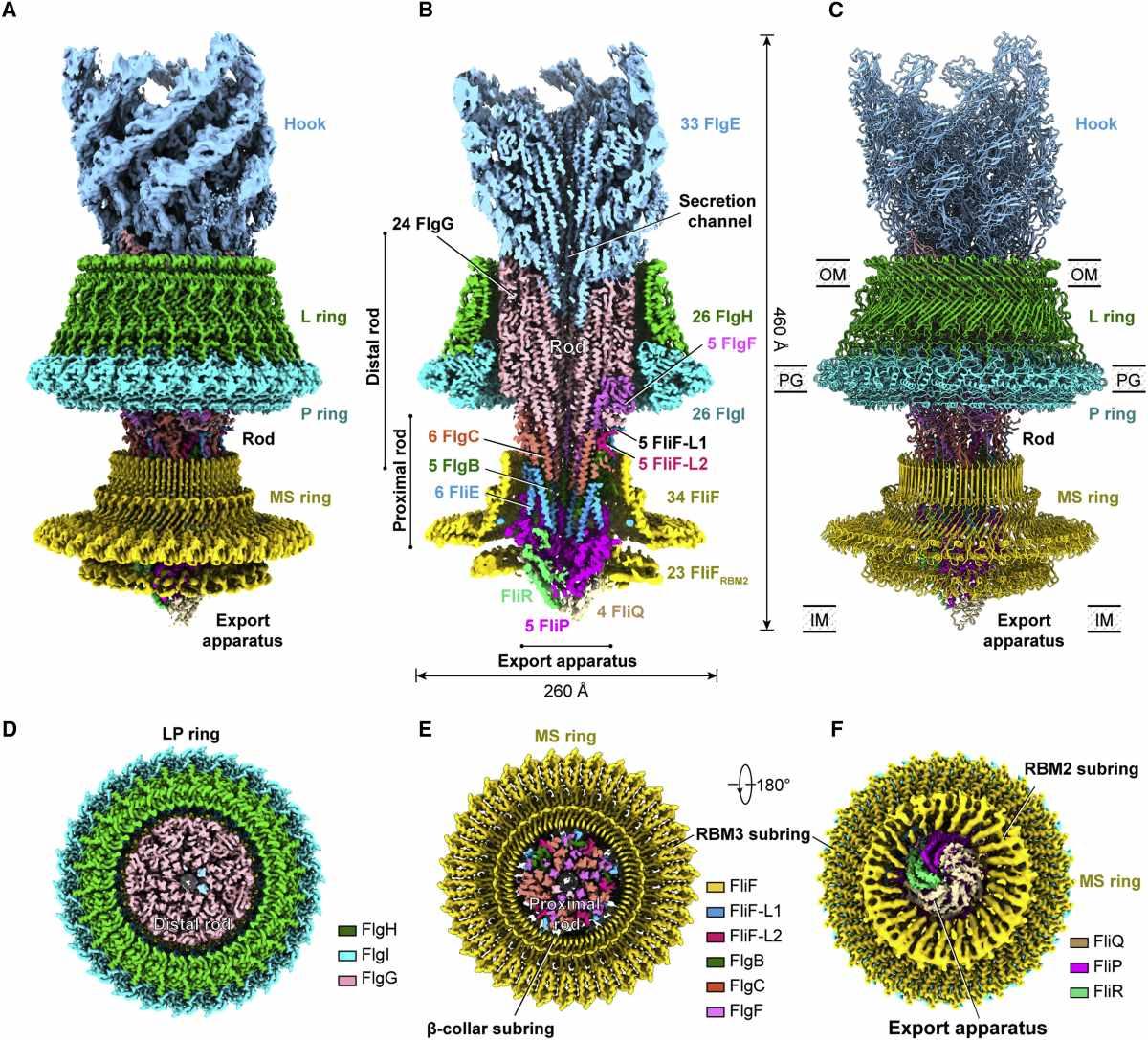
(A) Обединена карта на плътността на комплекса на флагеларен мотор-кука след локални уточнения. Картата е получена чрез монтиране на картите на плътност с висока разделителна способност на шест локално прецизирани региона (LP пръстен, кука, прът, експортен апарат и β-яка-RBM3 и RBM2 подпръстени на MS пръстена) в глобално прецизирания 3.9- Карта с Å-резолюция на комплекса мотор-кука.
(B) Изглед на напречно сечение на обединената карта на плътността на комплекса мотор-кука. Номерата на субединиците на компонентите са обозначени.
(C) Цялостната структура на флагеларния комплекс мотор-кука.
(D–F) Изгледи на напречно сечение на L пръстена (D) и MS пръстена (E) и изглед отдолу (F) на обединената карта на плътността на комплекса мотор-кука.
Куката и пръстените L, P и MS са оцветени съответно в светло синьо, зелено, циан и жълто. OM, външна мембрана; IM, вътрешна мембран
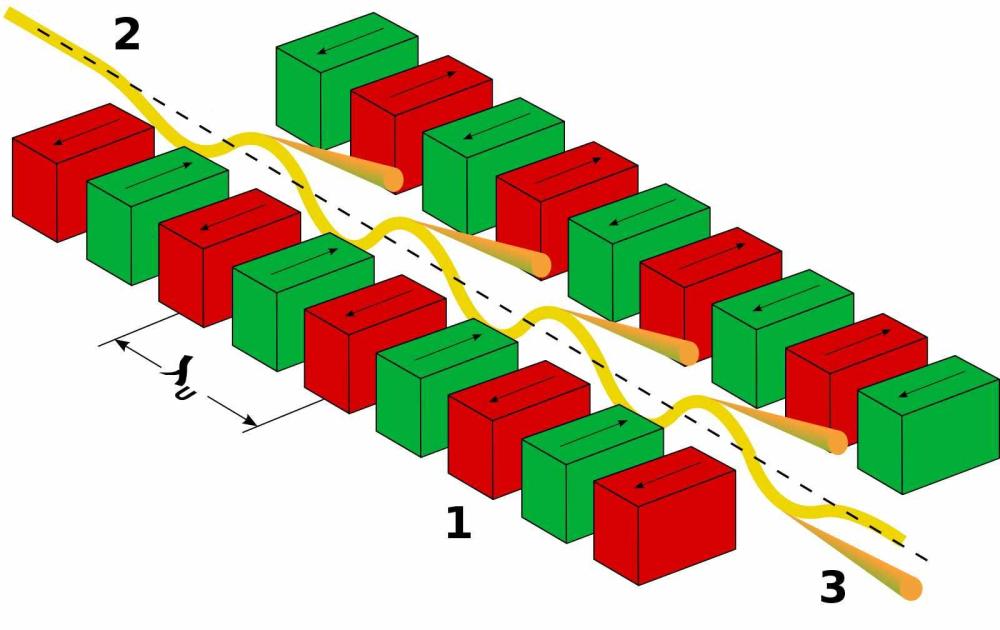
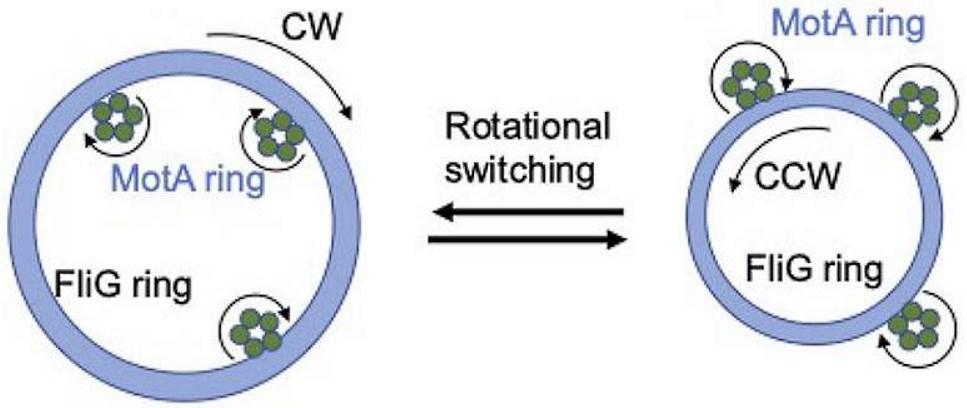
Всяка статорна единица се състои от два трансмембранни протеина, наречени MotA и MotB в E. coli и B. Burgdorferi. MotB е димер, заобиколен от MotA пентамерен пръстен с диаметър ~ 5−7,5 nm. Въз основа на нова структурна информация е установено, че MotA пентамерният пръстен се върти около фиксирания MotB димер. Следователно, всичките пет MotA субединици участват в генерирането на въртящ момент, независимо от посоката на въртене на ротора, а силовият удар се генерира директно от въртеливото движение на самите MotA субединици, вместо от техните вътрешни конформационни промени. Въпреки, че статорите са наречени заради тяхната „стационарна“ роля като невъртящ се компонент на двигателя, съставът на статорните единици, заобикалящи двигателя, е силно динамичен. Взаимодействието между MotA и протеините FliM на С-пръстена предизвиква въртеливо движение на ротора. По време на един силов удар пентамерният пръстен MotA се завърта за един период (2π/5). Всеки ход с пълна мощност се състои от два полутакта, всеки от които се захранва от протон, преминаващ последователно през един от двата йонни канала. По време на всеки половин ход, остатъкът от акцептора на протони (D32 за E. coli) върху съответния MotB мономер се протонира, което задвижва въртенето на MotA пръстена. В същото време протонираният MotB претърпява конформационна промяна, при която остатъкът на протонния акцептор се променя от позиция, обърната към периплазмата, към позиция, обърната към цитоплазмата, от която протонът може да бъде освободен в цитоплазмата.
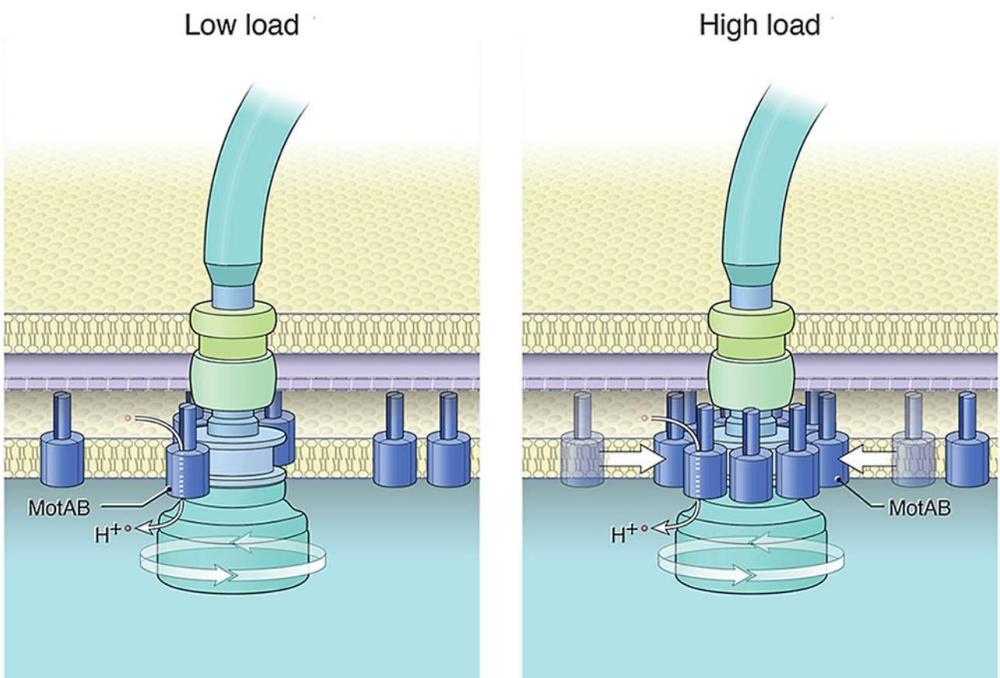
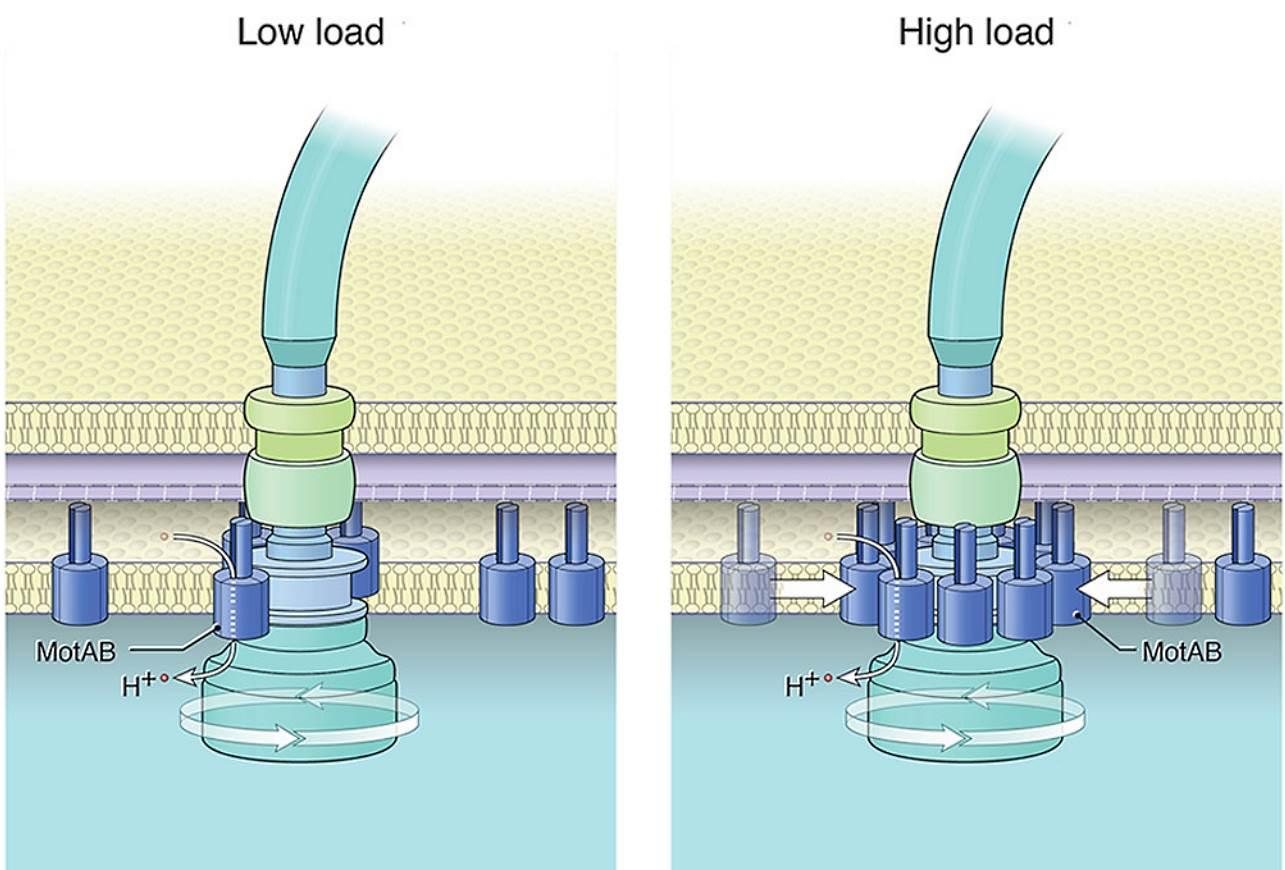
Докато бактериалните клетки се движат през хетерогенни среди, те срещат различни течности с различен вискозитет. По този начин, при променящи се условия, бактериите изпитват различни нива на вискозно съпротивление (или механично натоварване) върху техните флагели. В по-вискозни среди двигателите трябва да генерират повече сила, за да въртят камшичетата и го правят чрез набиране на статорни единици към двигателя по начин, зависим от натоварването. Експериментално е доказано, че статорните единици в двигателя варират от 0 до 17 (моторът на C. jejuni) в зависимост от механичното натоварване на двигателя. Когато работят 11 единици (Escherichia coli) за генериране на въртящ момент, това съответства на максимален въртящ момент от ∼200 pN⋅nm на всяка статорна единица.
С увеличаване на натоварването, скоростта на ротора бързо намалява до нула, докато скоростта на статорните единици се доближава до постоянна скорост (намалена, но все още ограничена). Въртенето на статора и ротора са синхронизирани при ниско натоварване т.е. няма приплъзване между двата въртящи се пръстена. Приплъзването между двата пръстена обаче се увеличава с натоварването. Близо до спиране скоростта на ротора изчезва, докато статорът все още се върти с крайна скорост. Наличието на приплъзване статор-ротор зависи от силата на съединителя, която е пропорционална на притискането статор-ротор (MotA-FliG).
В обобщение, статорните единици MotA представляват микромотори, които се въртят активно от протонната движеща сила. Те предават въртеливо движение на ротора чрез директен контакт. По този начин ротора получава въртеливо движение с което задвижва централния вал в края на който е флагеларната нишка-камшика.
Посоката на въртене се контролира от сложна хемотактична система. В сигналния път CheY се фосфорилира от CheA киназа, след което фосфорилираният CheY се свързва с FliM протеина в С-пръстена и индуцира конформационни промени, които променят взаимодействието статор-въртене и причиняват превключване. CheX е единствената CheY-P фосфатаза, идентифицирана в генома на B. Burgdorferi .
Като пример за „двукомпонентна” регулаторна система, фосфорилираният CheA отдава фосфат от фосфорил-хистидин на аспартатен остатък на регулаторния протеин CheY на реакцията. Фосфорилираният CheY (CheY-P) директно свързва флагеларния роторен протеин FliM, за да благоприятства въртенето по посока на часовниковата стрелка (CW) на камшичетата, насърчавайки разглобяването на флагеларния сноп и преобръщане на клетките. Когато CheA се деактивира от атрактанта, намаляването на CheY-P благоприятства въртенето обратно на часовниковата стрелка (CCW) на ротора.
Казано накратко, микромоторите на статорния комплекс се въртят само в една посока (CW) и предават въртеливо движение по външната страна на ротора, така той се върти обратно на часовата стрелка (CCW). След като ензима CheA киназа фосфорилира протеините CheY те се свързват с роторните протеини FliM на С-пръстена. Това свързване предизвиква конформационни промени в FliM, които водят до увеличаване диаметъра на С-пръстена от 55 nm на 63 nm и микромоторите започват да контакуват по вътрешната му страна предизвиквайки смяна на посоката на въртене.
Тук има видео от което се добива ясна представа за целия механизъм по смяна на посоката на въртене:
https://static-content.springer.com/esm/art%3A10.1038%2Fs41594-020-0497-2/MediaObjects/41594_2020_497_MOESM9_ESM.mp4
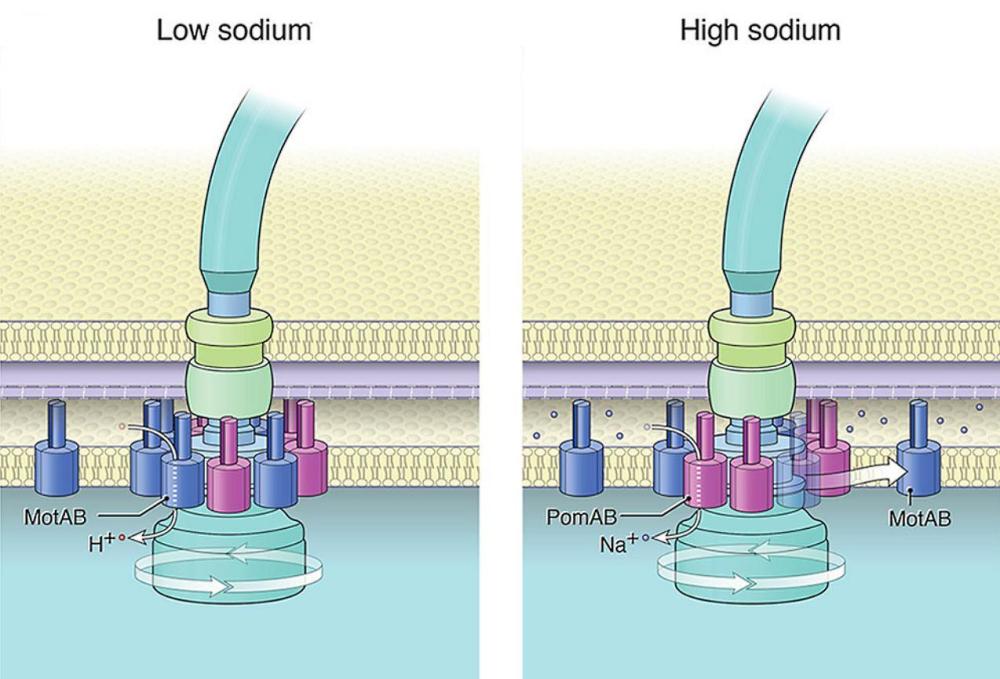
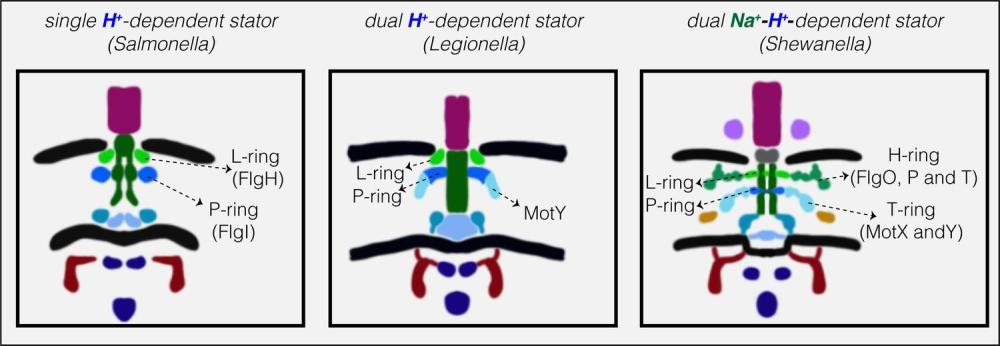
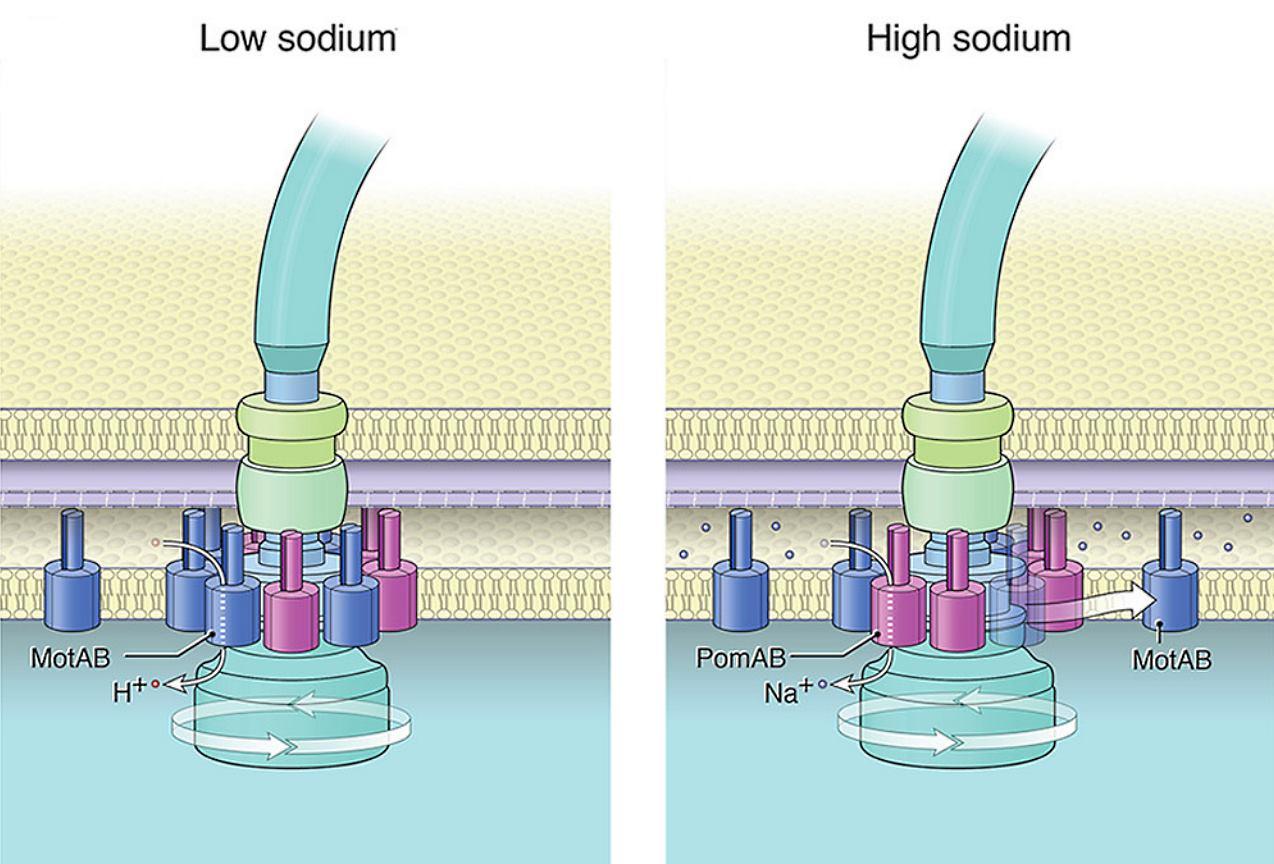
Някои бактерии съдържат както задвижвани от протони MotAB статори, така и задвижвани от натриеви йони PomAB статори. Работата с S. oneidensis показа, че броят на MotAB статорите, включени в двигателя, намалява с увеличаване на концентрацията на натриевите йони.
Динамиката на статора става по-сложна при видове с множество статорни системи, някои от които се задвижват от различни йони. Протон-зависимите статори захранват повечето флагели, но двигателите при няколко вида (особено тези в морска среда) зависят вместо това от натриевия поток. За да регулират двигателната функция в тези по-сложни системи, бактериите трябва да избират и да сменят типовете статори. Тези са факторите, управляващи разположението и обмена на статорните единици.
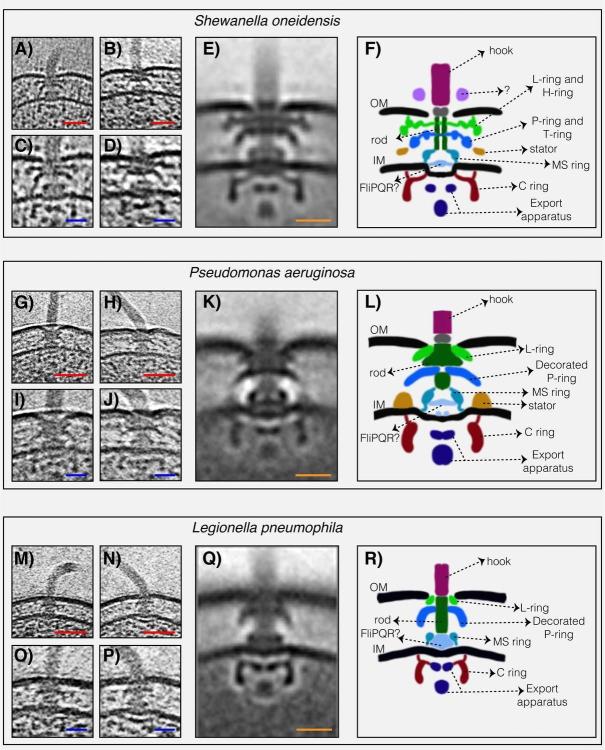
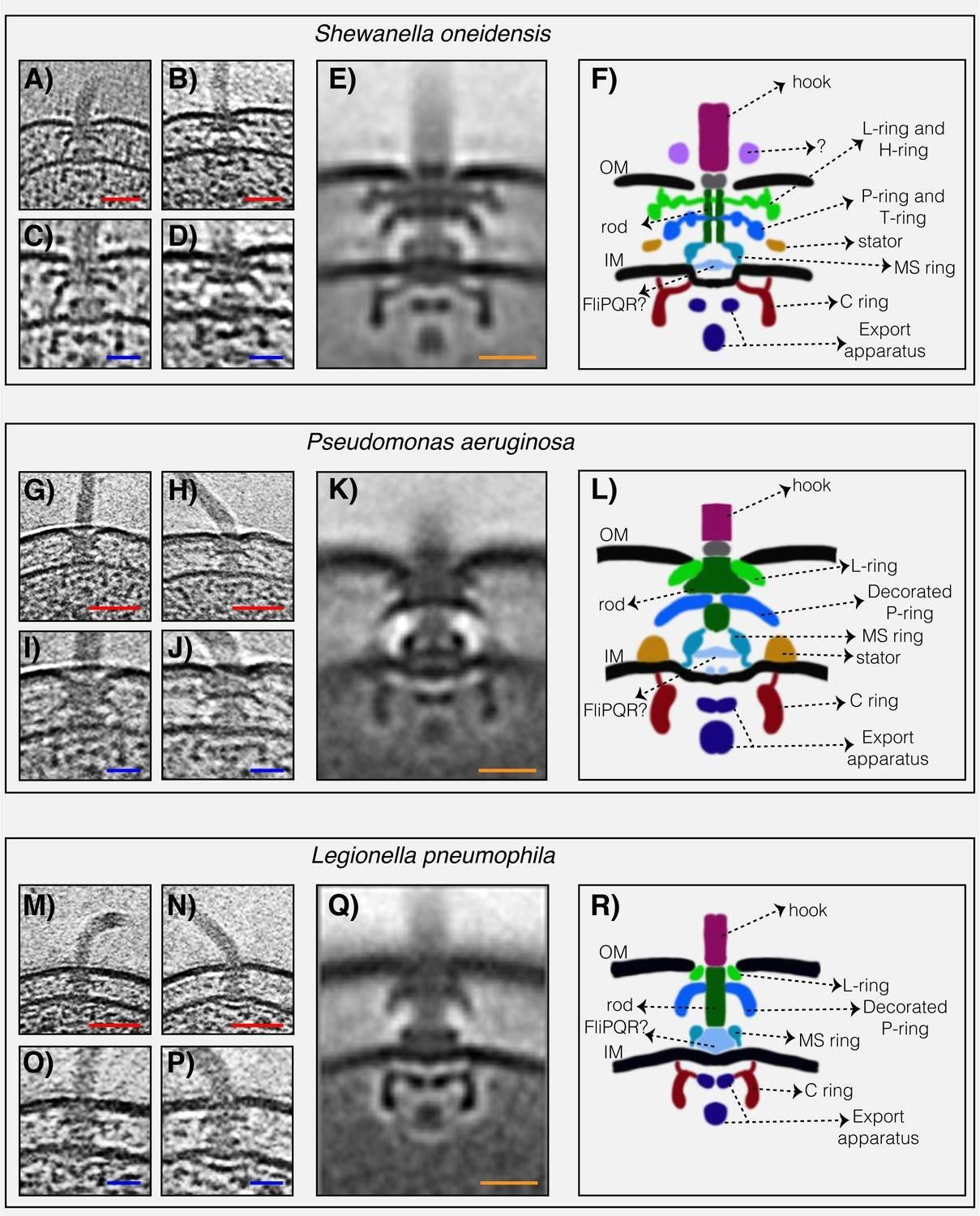
Структурите на три флагеларни мотора на Gammaproteobacteria с двоен статор, разкрити от ECT. ( A и B ) срезове през Shewanella oneidensis MR-1 електронни крио-томограми, показващи единични полярни флагели. ( C и D ) увеличени изгледи на резените, показани в ( A ) и ( B ), подчертаващи флагеларните двигатели. ( E ) централен срез през субтомограма средна на флагеларния двигател на S. oneidensis MR-1. ( F ) схематично представяне на средната субтомограма, показана в ( E ), с етикетирани основните части на двигателя. ( G–L ) флагеларен двигател на Pseudomonas aeruginosa . Панелите следват същата схема като в ( A–F ) по-горе. ( М–Р) флагеларен двигател на Legionella pneumophila . Панелите следват същата схема, както по-горе. Скала 50 nm (червена) и 20 nm (синя и оранжева).
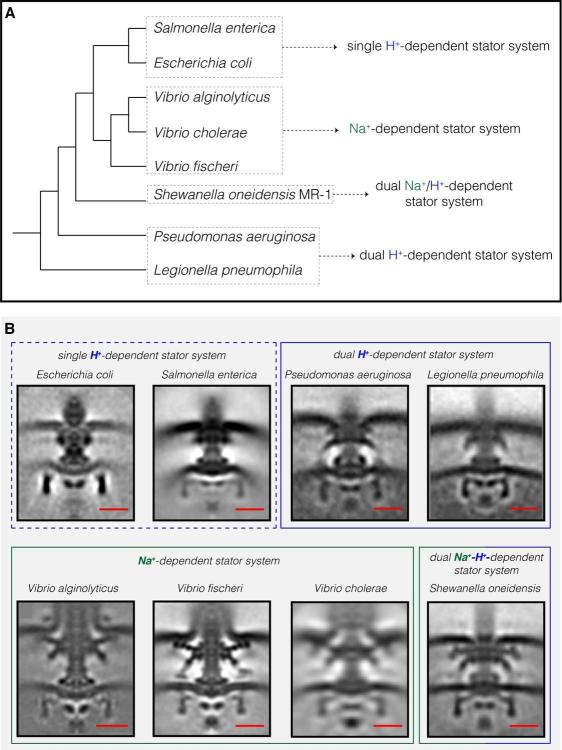
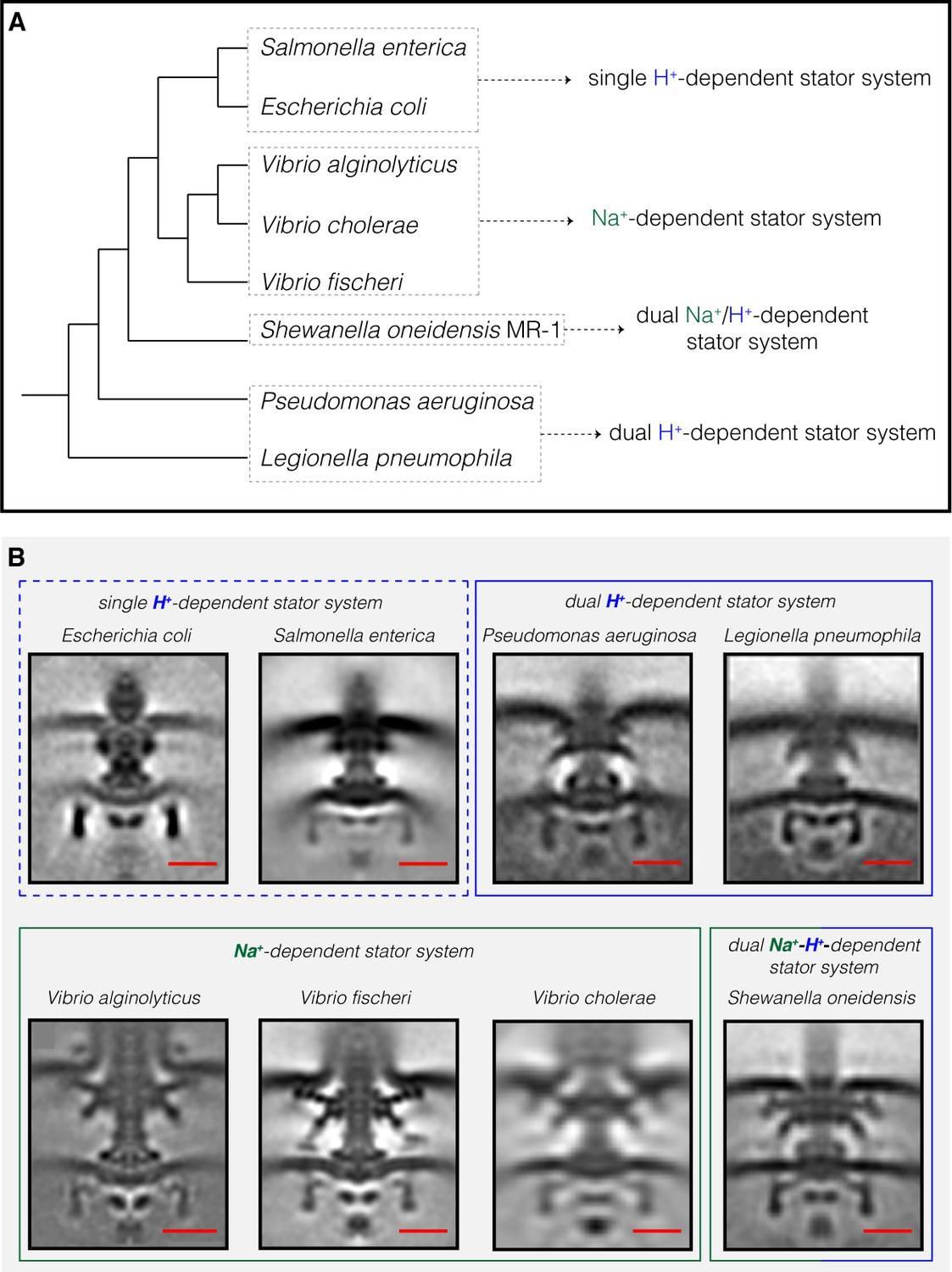
Компилация от всички флагеларни двигатели на Gammaproteobacteria, изобразени до момента от ECT. ( A ) Филогенетично дърво на осемте вида Gammaproteobacteria с налични ECT структури на техните флагеларни двигатели. Това дърво е направено въз основа на ( Williams et al., 2010 ). ( B ) Показани са централни срезове на средните субтомограми за осемте флагеларни мотора на Gammaproteobacteria, разкрити чрез ECT, включително трите структури, разрешени в това изследване ( P. aeruginosa, L. pneumophila и S. oneidensis ). Двигателите се класифицират въз основа на тяхната статорна система: единично H + -задвижване (пунктирана синя кутия), двойно H + -задвижване (синя кутия), Na + -задвижване (зелено поле) или двойно Na + -H + -задвижване (зелено - синя кутия).E. coli EMDB 5311, S. enterica EMDB 3154, V. fischeri EMDB 3155, V. cholerae EMDB 5308, V. alginolyticus
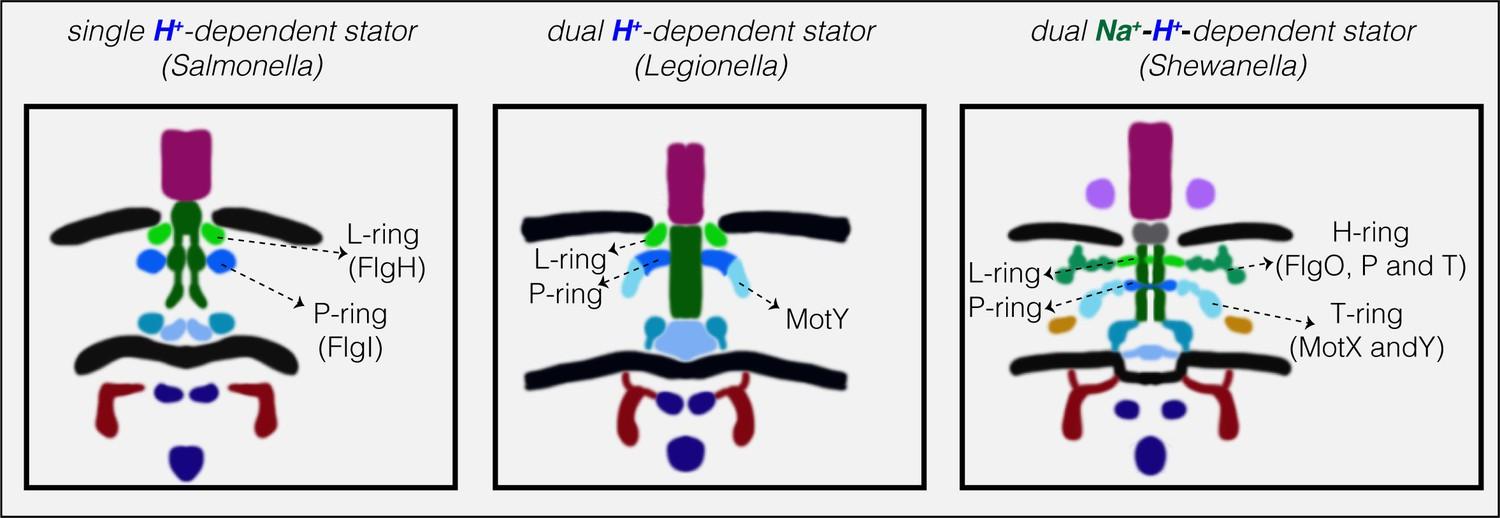
Нискоскоростните двигатели с единични H + -статорни системи имат само P- и L-пръстени, докато високоскоростните двигатели, използващи Na + , имат два допълнителни периплазмени пръстена, T- и H-пръстени.
Материали:
https://en.wikipedia.org/wiki/Flagellum
Динамичната йонна движеща сила, захранваща бактериалния флагеларен двигател
https://www.frontiersin.org/articles/10.3389/fmicb.2021.659464/full
Конструктивна основа на монтажа и предаването на въртящия момент на бактериалния флагеларен двигател
https://www.sciencedirect.com/science/article/pii/S009286742100430X?via%3Dihub
Механика на генериране на въртящ момент в бактериалния флагеларен двигател
https://www.pnas.org/doi/full/10.1073/pnas.1501734112
Моделиране на бактериален флагеларен двигател с нова информация за структурата: Ротационна динамика на два взаимодействащи протеинови нано-пръстена
https://www.frontiersin.org/articles/10.3389/fmicb.2022.866141/full
3D cryo-EM изобразяване на бактериални камшичета: Нови структурни и механични прозрения в клетъчната подвижност
https://www.sciencedirect.com/science/article/pii/S0021925822005464?via%3Dihub
Пропулсивни наномашини: конвергентната еволюция на archaella, flagella и cilia
https://academic.oup.com/femsre/article/44/3/253/5800988
Бактерии, усъвършенствайте двигателите си: Динамиката на статора регулира подвижността на флагелата
https://journals.asm.org/doi/10.1128/JB.00088-17
Нативният флагеларен MS пръстен се формира от 34 субединици с 23-кратни и 11-кратни субсиметрии
https://www.nature.com/articles/s41467-021-24507-9
Нов спин на флагеларното въртене
https://www.nature.com/articles/s41564-020-00820-9
Различни бактериални флагеларни двигатели с висок въртящ момент сглобяват по-широки статорни пръстени, използвайки запазено протеиново скеле
https://www.pnas.org/doi/full/10.1073/pnas.1518952113
Наличието и отсъствието на периплазмени пръстени в бактериалните флагеларни двигатели корелира с типа на статора
https://elifesciences.org/articles/43487
Сглобяване и динамика на бактериалния флагелум
https://www.annualreviews.org/doi/10.1146/annurev-micro-090816-093411
Структурно разнообразие на бактериални флагеларни двигатели
https://www.embopress.org/doi/full/10.1038/emboj.2011.186
Архитектура и монтаж на периплазмен флагел
https://journals.asm.org/doi/10.1128/microbiolspec.PSIB-0030-2019?url_ver=Z39.88-2003&rfr_id=ori:rid:crossref.org&rfr_dat=cr_pub%20%200pubmed
Поетапно формиране на бактериалната флагеларна система
https://www.pnas.org/doi/full/10.1073/pnas.0700266104
Ротационният двигател на бактериалните флагели
https://www.annualreviews.org/doi/10.1146/annurev.biochem.72.121801.161737